|
BT53A
|
| Fundamentos microbiológicos
de la ingeniería ambiental |
Leandro Herrera Z., Ph.D.
Laboratorio de Procesos Ambientales (INPRODES)
Departamento de Ingeniería Química
Facultad de Ciencias Físicas y Matemáticas
Universidad de Chile
Beauchef 861, Santiago, Chile
leherrer@cec.uchile.cl
http://tamarugo.cec.uchile.cl
678.4163 |
La actividad industrial del pasado siglo
tuvo un serio efecto acumulativo sobre el medio ambiente.
De hecho, el ritmo de producción
actual ya no es factible si utilizamos las mismas tecnologías que
producjeron el impacto ambiental acumulado.
Diversos efectos sociales:
-
actividades netamente contestatarias y
activistas
-
surguimiento de nuevas teorías
y enfoques económicos
-
efervesencia de desarrollo de "normas"
y leyes
-
modificación
de las normas de diseño de cualquier proceso industrial o proyecto
económico
¿Cómo los nuevos proyectos
no hipotecan el ambiente ni los recursos de futuras generaciones?
Nuestro Problema: comprender algunos
elementos básicos que permiten concebir los proyectos de ingeniería
en el contexto de un mundo que ha optado por cautelar el ambiente y los
recursos naturales en aras de la continuidad del desarrollo de la vida
en el futuro.
¿Cómo opera una industria
de transformación de materiales para la sociedad?
La
figura 1.1.1 contiene un boceto general de una planta industrial cualquiera,
vista como un diagrama en bloque (también se les llama caja negra
en términos coloquiales), que permite discutir el balance
global del proceso en cuestión.
Los insumos pueden ingresar
en cualquier fase:
-
sólida (indicada por Fse
en la figura, para abreviar Flujo de Entrada de Sólidos);
-
líquida (indicada por FLe,
Flujo Líquido de Entrada);
-
gaseosa (que no se ha indicado en la entrada
de esta planta, aún cuando es típico que se consuma aire),
-
servicios de energía (indicado
por FHe por Flujo de Energía de Entrada, etc.).
Similarmente los productos
pueden ocurrir en cualquier fase; en las salidas si se ha indicado el caudal
de gas puesto que muchas plantas industriales producen emisiones
(salidas en fase gas que no forman parte del
producto final) en la forma de gases de combustión; olores; polvo;
etc..
Cualquiera que sean las transformaciones
que se produzcan sobre la materia prima, esta estará sujeta a reacciones
(de síntesis o de separación); a procesos de transporte (por
ejemplo, llevar sólidos de un punto a otro mediante un caudal de
agua); a procesos termodinámicos, etc.
Es poco probable que todos los elementos
de los insumos participen del producto; más bien, es común
que los productos formen sólo una fracción (másica)
pequeña de los insumos totales.
En balance, naturalmente, sobrarán
productos secundarios que de no ser atendidos específicamente y
si este descarte ocurre al medio ambiente, en cualquier fase, y si su concentración,
volumen o masa excede los valores definidos por la ley (que es una expresión
de un acuerdo social), entonces esa descarga es contaminante.
Sabemos de los efectos nocivos de los
contaminantes,
en primer lugar, por la incidencia que tienen sobre la salud
humana, tanto de los trabajadores que participan directamente de
la actividad como de los habitantes y de las especies vivas en general.
Secundariamente, por la perdida
de calidad del suelo, de las aguas
y de la atmósfera circundantes a la
actividad industrial en cuestión.
En tercer lugar, por la disminución
cuantitativa de los recursos naturales no
renovables (si bien Chile aún no dispone de Cuentas Ambientales,
de modo que esta disminución es más bien de tipo cualitativo).
Es decir, el primer
indicador es biológico, el segundo
es físico químico y los tres comparten la componente
económica; en particular, la mala salud significa costos
de tratamiento y ausentismo laboral; los suelos reducen su capacidad de
producción
agrícola; los recursos no renovables (cobre, por ejemplo) se reducen
progresivamente.
| La evaluación ambiental, por
ende, podría ser una evaluación técnico económica
que incorpora la factibilidad de la no contaminación. |
En el largo plazo, el efecto de una actividad
contaminante suele extenderse significativamente más allá
del sector geográfico de la actividad industrial, debido a los mecanismos
de transporte propios de la biosfera (no se puede pedir pasaporte
a gases o líquidos contaminados que cruzan la frontera de un país
a otro).
En cuanto a la reacción de la
sociedad, post-constatación de la contaminación, se ha reglamentado
la cuantía aceptable de aquellos elementos que se sabe (o
se sospecha razonablemente) que tienen un impacto
ambiental importante; la propia estimación del impacto ambiental
de una actividad es sujeto de una legislación y metodología
específicas.
El nuevo marco legal aparece como nuevas
restricciones del diseño industrial, con lo que se ha producido
un reemplazo de tecnologías tradicionales -hoy llamadas sucias-
por otras más
limpias y una
serie de proposiciones de modificación de las normas de diseño
de nuevos proyectos de inversión, como por ejemplo, la metodología
de emisión cero, la metodología
de reducción de la contaminación y otras más.
Simultáneamente,
se hace necesario proveer soluciones para amortiguar
el
impacto ambiental de procesos ya existentes, es decir, diseñar procesos
para el tratamiento de los residuos. En este caso, nos ocupará el
tratamiento de los residuos industriales líquidos,
ya que sólo los líquidos pueden
interactuar con los sistemas biológicos; las fases sólida
y gaseosa aparecerán sólo en tanto sus compuestos puedan
entrar en solución (por ejemplo, el SO2 es nocivo porque
al disolverse y reaccionar en la humedad de las vías respiratorias
genera ácido sulfuroso que nos daña los tejidos).
Dado que el impacto ambiental de un
proceso está centrado, entre otros aspectos, en la vida
humana, es necesario entender mínimamente la naturaleza
del impacto biológico, e introducir aquellos aspectos en
que se basan las tecnologías biológicas de tratamiento de
residuos, tanto industriales (RILES) como urbanos (Aguas Servidas).
Para el estudio del tema será
necesario:
-
introducir breves aspectos de microbiología
fundamental
-
revisar algunos aspectos de salud
humana, desde el punto de vista epidemiológico;
-
aspectos de microbiología
aplicada
-
ecología
básica
-
fundamentos de diseño
de procesos biológicos
-
revisar las diversas teorías
filosóficas que recogen los antecedentes históricos
de la relación que ha establecido la humanidad con el planeta y
su naturaleza, puesto que se puede observar un resurgimiento de viejas
ideas (la Tierra está viva,
por ejemplo) para explicar y reaccionar frente a la era ambiental.
Elementos
de Microbiología Básica
Clasificación de Microorganismos
La microbiología estudia las formas de vida más
pequeñas (microbios), de tamaño
microscópico; es decir, se requiere un microscopio para verlos.
En términos históricos, entre los primeros
esfuerzos para clasificar, ordenar
y caracterizar nuestra comprensión
del mundo y sus más obvios elementos, se habló de los reinos:
-
animal,
-
vegetal y
-
mineral.
En esta clasificación no quedó claro, posteriormente,
a que grupo adscribir los microorganismos
que se "descubrieron" gracias a la invención del microscopio. Los
microorganismos se subclasificaron, después, en:
-
algas,
-
protozoos,
-
hongos,
-
bacterias y, más tarde,
-
virus.
Haeckel propuso un nuevo reino, el protista
para clasificarlos (los virus fueron descubiertos más tarde). El
reino protista hubo de ser subdividido -al
avanzar el conocimiento sobre las estructuras celulares- en los protistas
superiores llamados eucariotes
e inferiores, llamados
procariotes.
Los eucariotes son microorganismos que tienen el material
nuclear separado del citoplasma por una membrana nuclear, mientras que
los procariotes no poseen membrana nuclear, manteniendo el material nuclear
(en particular, el material genético, ADN) en contacto con todos
los otros componentes de la célula (en el citoplasma).
La consecuencia directa de estas diferencias en la
protección del material nuclear es la mayor mutabilidad (cambio)
de los procariotes versus la mayor estabilidad genética de los eucariotes,
cuyo material genético está más estructurado y más
protegido.
Según se pudo observar más tarde, sólo
las bacterias y las entonces llamadas algas azul-verde presentan
características procariotes, es decir,
no tienen membrana nuclear.
Hoy en día, las algas azul-verde
son consideradas bacterias y se las llama,
en particular, las cianobacterias.
De este modo, los términos bacteria
y procariote son sinónimos.
Los hongos, las algas y los protozoos
constituyen los protistas eucariotes. En síntesis, los microorganismos
incluyen:
-
los protistas eucariotes,
-
los protistas procariotes (bacterias)
y
-
los virus.
Al discutir la participación de los microorganismos
y su interrelación con los efluentes de procesos industriales, se
debería revisar la posible contribución de cada tipo de microorganismo
tanto como de las interrelaciones entre ellos.
Desde un punto de vista global, las bacterias juegan
un rol más importante que todos los otros microorganismos, ya que
forman la base de los ciclos de reciclaje
de nutrientes y elementos del ecosistema (sin desmerecer la base energética
de los orgánicos del planeta, que está en los fotosintéticos
-algas y cianobacterias- que sintetizan orgánicos a partir del CO2
y de luz solar).
Las bacterias más estudiadas han sido las patógenas
(particularmente para los humanos). Sin embargo, muchas otras bacterias
no patógenas son del mayor interés tecnológico, por
ejemplo aquellas que participan en depuración
de aguas servidas o industriales; depuración natural en ríos,
esteros o lagos; descomposición de
orgánicos en el suelo; relleno sanitario (metanogénesis)
y aquellas que forman la base de los procesos biotecnológicos que
utilizan la ingeniería (o manipulación) genética.
Los virus, mucho más pequeños que las
bacterias (virus: 0,01 a 0,25 m m, bacterias:
1 a 4 µm), basan su ciclo vital en la maquinaria celular de alguna
célula receptora. Existen viruces que invaden las células
de todos los otros seres, tanto bacterias como plantas y animales; pero
cada
virus es más bien específico a su célula receptora.
Los virus suelen considerarse en la
frontera de lo
vivo ya que no es simple su adscripción, al no disponer de
una maquinaria que les permita su propia síntesis. La figura
2.1.1 contiene bocetos de una bacteria y un virus típicos, a
fin de contrastar sus diferencias morfológicas y de tamaño.
Las algas (y las cianobacterias) forman la maquinaria
necesaria para la síntesis orgánica
a partir de la energía solar y carbono
inorgánico (el dióxido de carbono, en particular).
Son verdaderos transductores
de energía solar (luz y calor) a energía de enlaces
químicos.
Toda la cadena alimenticia del globo terráqueo
descansa sobre la fotosíntesis (si bien se conocen algunos casos
de utilización de potenciales de óxido reducción de
elementos inorgánicos), de modo que su importancia es del todo evidente.
En cuanto a la posible utilización de algas
fotosintéticas en aplicaciones industriales,
esta se encuentra limitada por la dificultad de construir
grandes reactores permeables a la luz, solar o artificial, que permita
el manejo de una explotación a gran escala. Sin embargo, existen
consideraciones de alta escala a considerar: en primer lugar, las aguas
que reciben excesos de nutrientes suelen alojar una cantidad muy grande
de algas, a tal punto que su crecimiento excede los límites de densidad
natural y se agota el sustrato, problema conocido como el florecimiento
de algas; además, las algas son del mayor interés
para el tratamiento de aguas servidas en lagunas (o pozos) ya que permiten
mantener condiciones de oxígeno disuelto por sobre el valor de saturación
en agua en la fracción superior (menos profunda), donde penetra
la luz solar.
Los hongos
son seres que existen tanto en forma unicelular como multicelular (cenocíticos),
no-fotosintéticos, que pueden sobrevivir en las más diversas
condiciones de pH (acidez o alcalinidad). Su participación en aplicaciones
industriales es amplia y tradicional, conocida desde hace muchísimos
años, aun antes de la revolución industrial (baste mencionar
la levadura del pan y la cerveza). En cuanto al tratamiento de aguas, son
importantes tanto en los tratamientos de aguas residuales industriales
como urbanas, además de la compostación de sólidos.
Muchas plantas (orquídeas,
por ejemplo, mycorrhizal) no pueden vivir sin una estrecha relación
con hongos simbióticos.
Los protozoos
son más grandes que las bacterias (alrededor de un orden de magnitud
mayor) y suelen participar en procesos biológicos de purificación.
Se pueden considerar como protistas otros microorganismos
del tipo crustáceo, los putrefactores y otros que, en último
análisis, caen dentro de la clase de los pequeños animales
más que en alguna de las anteriores.
La figura
2.1.2 contiene un boceto, sumamente simplificado, de una bacteria.
Según se ha dicho, la diferencia principal entre una bacteria (procariote)
y un eucariote es la presencia de una membrana nuclear en los últimos,
mientras que los eucariotes sólo tienen diferenciada una región
nuclear (que contiene el ADN). La figura destaca, además, la presencia
de un pared y de una membrana celulares, amén de corpúsculos
(ribosomas) que participan de la decodificación del material genético
para la construcción de proteínas.
-
Bacterias
Las bacterias (bacteria plural; bacterium singular) son
seres unicelulares microscópicos, de formas extraordinariamente
variadas y que se encuentran prácticamente en todos los habitat
de la biosfera. La forma puede ser de una de tres estructuras geométricas
generales: esférica (cocci, coccus), cilíndrica o bastón
(bacilli, bacilus) y espiral (spirila, spirilum). Además, muchas
especies tienden a vivir asociadas en grupos de a pares, en cadenas, en
pelotones, etc. En la tabla 2.2.1 se han seleccionado algunas bacterias
de interés para la temática industrial.
Las bacterias contienen una serie de estructuras (evidentemente
aun más pequeñas que la propia célula) de propósito
específico: genes en cromosomas, ribosomas, membranas, entre otras.
Todas estas sub-estructuras se encuentran en suspensión en un medio
líquido, llamado el citoplasma, donde se disuelven, además,
las proteínas, azúcares, ácidos grasos, etc. que participan
del ciclo vital de la célula.
TABLA 2.1.1: Selección de algunas Bacterias
de interés industrial o epidemiológico
| Grupo |
Género |
Importancia
(característica) |
| Indicadores |
Escherichia
Enterobacter
Streptococcus
Clostridium |
Se les utiliza
como indicadores de contaminación de una fuente de agua, por contacto
con feces humanas. Se utilizan como indicadores aquellas bacterias
cuyo hábitat usual es principalmente humano. |
| Patógenos |
Salmonella
Shigella
Mycobacterium |
Fiebre tifoidea
Disentería
Tuberculosis |
| Nitrificadores |
Nitrobacter
Nitrosomonas |
Oxidan compuestos
inorgánicos de nitrógeno. |
| Denitrificadores |
Bacillus
Pseudomonas |
Reducen nitritos
y nitratos a nitrógeno molecular gaseoso o a óxido nitroso
gaseoso. |
| Bacterias
del decaimiento o de la degradación |
Pseudomonas
Flavobacterium
Zooglea
Clostridium
Micrococcus
Methanobacterium
Methanococcus
Methanosarcina |
Degradación
de orgánicos
Degradación de proteínas
Formación de flóculos
en lodos activados
Productor anaerobio de ácidos
grasos
Productor anaerobio de ácidos
grasos
Productor anaerobio de metano a partir
de ácidos grasos (útiles en reactores de metanogénesis). |
| Bacterias del
Azufre |
Thiobacillus
Desulfovibrio |
Oxidadora
de azufre y de hierro
Reductora anaerobia (azufre y otros,
corrosión) |
| Bacterias fotosintéticas |
Chlorobium
Chromatium |
ambos
géneros reducen sulfuros (S=) a azufre elemental. |
Ciclo de vida de las bacterias
El proceso de vida de una célula bacteriana puede
ser simplificado a una visión de acumulación de sus propios
componentes a partir de compuestos más simples que ella (el sustrato)
mediante un proceso conocido como metabolismo celular. El concepto
de sustrato engloba todos los requerimientos nutricionales y físicos
de la célula y, naturalmente, dependen de la especie en particular.
Una de las variables manipulables más importantes es la temperatura
y, en este sentido se debe tener en cuenta que hay bacterias que se pueden
desarrollar a temperaturas bajo 0ºC y otras crecen óptimamente
a temperaturas del orden de los 70ºC y otras pueden tolerar, incluso,
los 99ºC. Sin duda, lo más típico es el rango desde
la temperatura ambiente hasta los 70ºC.
Desde el punto de vista de su forma
nutricional, se distinguen dos grandes grupos: los heterótrofos
y los autótrofos. Las bacterias
heterótrofas obtienen tanto su energía como su carbono a
partir de moléculas orgánicas. Los autótrofos, en
cambio, obtienen el carbono para su desarrollo desde el dióxido
de carbono, CO2, y la energía a partir de compuestos
inorgánicos (sales que resultan oxidadas) o de la luz (en principio,
solar). Si el autótrofo del caso utiliza luz, se le suele llamar
fotoautótrofo mientras que si
utiliza sales se le llama quimioautótrofo
(litoautótropho).
El soporte a la vida celular incluye no sólo
el carbono y la energía; además se deben contemplar fuentes
de potasio, fosfato, nitrógeno, azufre, metales y otros elementos
en bajas concentraciones y que dependen de cada especie. Dos medios de
cultivo típicos se han resumido en la tabla 2 (en particular, el
medio de cultivo de la bacteria quimioautótrofa Thiobacillus
ferroxidans, de amplio uso en la lixiviación bacteriana
de minerales de cobre u otros y de la bacteria anaerobia heterótrofa
Desufovibrio
desulfuricans, utilizada para la reducción de sulfatos
en aguas de mina).
Desde otro punto de vista, considerando que las variables
físicas de mayor interés son la temperatura, el pH y la concentración
de gases atmosféricos en la fase líquida, las bacterias se
clasifican en grupos que dependen de estas variables. Así, respecto
de la temperatura, se habla de psicrófilos
para las bacterias capaces de crecer a temperaturas bajo 0ºC hasta
los 25ºC; de
mesófilos
para aquellos capaces de crecer en el rango de los 25 a los 40 ºC
y de termófilos para aquellos
capaces de crecer entre 40 y 100 ºC. Desde el punto de vista del pH
se definen como bacterias
acidófilas
aquellas que pueden vivir en condiciones de acidez extrema (pH 1 a 2).
Tabla 2.2.2: Ejemplos de
medios de cultivo para el desarrollo de bacterias
| Thiobacillus |
ferrooxidans |
Desulfovibrio
|
desulfuricans |
| Compuesto |
[g/L] |
Compuesto |
[g/L] |
| MgSO4×7H2O |
0,4 |
Na2SO4 |
3,0 |
| (NH4)2SO4 |
0,4 |
MgSO4 |
1,5 |
| K2HPO4×3H2O |
0,056 |
NH4Cl |
1,0 |
| |
|
K2HPO4×3H2O |
0,65 |
| |
|
CaCl2 |
0,08 |
| |
|
Fe(NH4)2(SO4)2×6H2O |
0,01 |
| |
|
Extracto de
levadura |
0,2 |
| |
|
Lactato, 85% |
5 mL |
| pH |
1,6 |
pH |
7,0 |
En cuanto a la clasificación de
acuerdo a los gases, son tres los gases de mayor importancia: el oxígeno,
el dióxido de carbono y el nitrógeno molecular. Aquellas
bacterias que crecen utilizando el oxígeno disuelto en su ciclo
respiratorio reciben el nombre de bacterias aerobias,
mientras que aquellas que crecen en ausencia de oxígeno son clasificadas
como
anaerobias. También se
habla de bacterias facultativas para
aquellas que pueden crecer tanto en ausencia como en presencia de oxígeno,
mientras que se llama obligadas a aquellas
bacterias que no tienen alternativa (una bacteria anaerobia
obligada es aquella que sólo puede desarrollar sus procesos
vitales en ausencia de oxígeno molecular disuelto). De menor ocurrencia,
se habla de bacterias microaerófilas
para englobar aquellas que pueden crecer con cantidades bajísimas
de oxígeno disuelto.
-
Desarrollo Celular Bacteriano
El metabolismo celular ocurre al interior
de la célula y lleva, como se describió anteriormente, a
la acumulación de componentes moleculares de la propia célula.
Esta acumulación (aumento de la concentración) de componentes
lleva a la elongación física de la bacteria. La elongación
de la bacteria y/o la acumulación de componentes llega a su cima
en el momento en que la célula inicia una división,
mediante una serie de pasos conocidos más por su evidencia visible
que por su regulación interna precisa. La primera evidencia es la
aparición de una cintura en el centro
de la célula y una distribución del material celular en dos
sectores. La cintura se sigue pronunciando hasta producir, en la práctica,
dos
nuevas células poco "elongadas" (se podría decir que
son dos células "delgadas" o "pequeñas", si bien estos términos
resultan muy difusos a nivel celular). La
figura
2.3.1 contiene un boceto de este proceso, enunciando sus etapas principales.
Este sistema de replicación celular es, naturalmente, más
conocido que la descripción recién expuesta, sin embargo,
desde el punto de vista de la observación empírica del aumento
del número de células en un sistema en que estas crecen,
la descripción es adecuada.
El tiempo que tome el ciclo desde el
nacimiento de una célula hasta la producción de dos nuevas
células es una función tanto de la propia célula como
de las condiciones ambientales en que se desarrolla. Sin embargo, una vez
garantizada la optimalidad del medio de desarrollo, la célula tiene
un tiempo de replicación que bien puede parecer una característica
intrínseca de esa especie. Es importante estar advertido de que
este tiempo de replicación mínimo puede variar notablemente
de especie en especie (por ejemplo, desde la veintena de minutos hasta
la decena de horas).
El tiempo medio que toma una cantidad
de células en duplicarse se conoce como tiempo
de replicación y es, en principio, el tiempo medio para
que una célula crezca y se divida en dos por fisión
binaria.
Según la descripción
anterior, en condiciones de abundancia de materiales de soporte (sustrato)
del metabolismo de una especie de bacterias, la tasa de crecimiento del
número de células en un medio dado, sería proporcional
al número de bacterias inicialmente presentes, es decir:
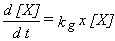
donde [X]: concentración
de células (número de bacterias por litro) a tiempo t;
kg:
constante de crecimiento de primer orden y d[X]/dt: tasa de crecimiento
celular (número de bacterias por litro y por unidad de tiempo).
La relación lineal se sostendrá mientras los sustratos sean
abundantes (por unidad de células) y mientras los productos finales
del metabolismo no interfieran con él mismo; es decir, en condiciones
ideales.
Diversas manipulaciones permiten, después
de integrar, llegar a cualquiera de las siguientes expresiones:
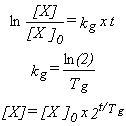
donde [X]0: concentración
de células a tiempo cero, Tg: tiempo de replicación.
De las ecuaciones integradas anteriores,
es claro que un gráfico logarítmico deberá mostrar
una recta al graficarse la concentración de células (en logaritmo)
versus el tiempo. Sin embargo, la visión recién presentada
es muy simplista, comparada con los resultados reales. La figura
2.3.2 muestra un perfil típico de un cultivo en lotes confinados
(batch), donde se puede observar que sólo una pequeña región
tiene un comportamiento lineal ascendente (marcada como fase logarítmica
o exponencial).
La fase anterior a la logarítmica
corresponde a la fase de latencia (lag) que es el tiempo que toma
a las células, recién iniciado el cultivo, para sintetizar
las enzimas necesarias y desarrollar sus componentes hasta tener suficiente
material como para iniciar la fisión binaria (otro nombre para proceso
de permanente crecimiento y división celular, basada en que de una
célula se obtienen dos); esta fase puede tener una duración
extensa o breve, según sea el estado fisiológico de las bacterias
iniciales, llamadas el inoculo. Existe
una transición gradual entre la fase de latencia y la logarítmica
que corresponde al momento en el cual más y más células
llegan al punto óptimo metabólico e inician el proceso de
síntesis y división a la más alta velocidad posible.
Posterior a la fase logarítmica,
se inicia la fase estacionaria, durante
la cual no hay un aumento importante en la concentración de células;
las razones son variadas y no están del todo claras pero parece
existir un "tope" de densidad de células que pueden existir en un
medio líquido. Por otra parte, puede corresponder al momento en
el cual se ha agotado algún nutriente pero aun existe sustrato energético
y carbónico que permiten la sobrevivencia celular, sin posibilitar
la división en nuevas células.
Finalmente, el cultivo envejecido y
sin sustrato inicia la fase de muerte
o de declinación, período en
el cual se observa una disminución del número de células
vivas. Nuevamente, las razones de la muerte celular no están del
todo dilucidadas pero es evidente que más de algún sustrato
se ha agotado; este agotamiento obliga a las bacterias a utilizar compuestos
energéticos almacenados a fin de sobrevivir hasta que estos compuestos
se agotan; además, se acumulan compuestos de desecho del metabolismo
que podrían resultar tóxicos para ellas mismas. Algunas bacterias
esporulan
(forman esporas) durante esta fase. Las esporas son partículas de
mayor densidad y menor tamaño que las bacterias originales pero
que contienen todo el material necesario para reconstruir la célula
una vez establecidas nuevamente las condiciones de cultivo.
En la figura se ha capturado, además,
la naturaleza del "ruido experimental" típicamente asociado a las
mediciones de densidad celular; se puede observar que los puntos experimentales
sólo están "cerca" de la curva que se podría trazar
entre los puntos. Habitualmente, la medición de densidad celular
es "ruidosa".
El objetivo de cualquier proceso biológico
de escala industrial, en general, no es sino la mantención de las
células en un estado permanente de crecimiento en la fase logarítmica.
Esto se consigue mediante reactores continuos, en los que entra sustrato
a un caudal idéntico al caudal de salida de productos (incluyendo
la biomasa).
Tal como es posible definir las condiciones
óptimas de desarrollo de las bacterias, es también posible
definir las condiciones más adversas para ellas. Existen diversas
razones por las que sería útil la más completa eliminación
de las bacterias de un líquido (por ejemplo, leche envasada, productos
alimenticios en general, agua potable, etc.).
Las técnicas
de eliminación de microorganismos son diversas y la que se
ocupe dependerá del propósito a la mano. Por ejemplo, un
líquido puede ser esterilizado,
mediante vapor de agua a 121ºC (a una presión de 15 psig) durante
unos 15 minutos, este tratamiento no sólo destruirá bacterias
sino que también esporas y hongos. A fin de evitar la verdadera
cocción que significa esta esterilización, se suele operar
con un ciclo de temperatura brusco, en el proceso conocido como UHT (Ultra
High Temperature) u otros procesos orientados a la pasteurización
más
que a la esterilización total. La esterilización destruye
las bacterias (y otros) mediante la denaturación
de las proteínas, principalmente.
Es fácil deducir la imposibilidad
de esterilizar un caudal de agua potable (por ejemplo de unos 4m3/seg
que está originalmente a unos 8 ºC). Por otra parte, puede
no ser necesario esterilizar el agua potable sino que puede bastar su desinfección,
es decir, eliminar los patógenos
(literalmente: agentes que producen enfermedades) acarreados por el agua.
La desinfección se suele conseguir mediante agentes oxidantes fuertes
y opera por disrupción de la membrana celular y por oxidación
de las proteínas y otros materiales citoplasmáticos. Los
oxidantes más típicos son el cloro y el ozono. Es prudente
también destacar que la legislación chilena exige que el
agua potable tenga un remanente de cloro incluso al llegar al punto de
consumo (las casas de una ciudad) a fin de garantizar que existe siempre
un potencial oxidante que impedirá el desarrollo de microorganismos.
Existen otros métodos de destrucción
de microorganismos, mediante radiación, Por ejemplo, la radiación
ultravioleta (longitud de onda entre 200 y 400 nm) destruye los enlaces
de los ácidos nucléicos y se la utiliza para desinfectar
las superficies de trabajo de instalaciones asépticas.
-
Virus
Los virus son, al menos, un orden de magnitud
más pequeños que las bacterias, cayendo en magnitudes típicas
desde 10 nm a 250 nm (1 nm= 1 nanómetro= 10-9 metro,
es decir, una millonésima de milímetro). En cambio, una bacteria
pequeña como la Salmonella
typhi (patógena responsable del tifus) mide unos 1000 nm,
o, en la nomenclatura más usual, 1 µm.
Desde un punto de vista fundamental,
es posible decir que la vida es una propiedad de los seres autopoyéticos,
que significa que se producen a sí mismos (por ejemplo, un animal
vivo que se rompe unas cuantas células de su piel, es decir una
herida, genera nuevamente dichas células de modo que se produce
a si mismo).
En virtud de dicha definición,
y tal como se mencionara más arriba, los virus plantean un problema
límite a la definición de lo vivo. De hecho, no contienen
sino ADN (o ARN) y la proteína de su envoltorio (cápsida)
y, por no contener ninguna de las sub-estructuras necesarias para la síntesis
ni para el metabolismo, no pueden producirse a si mismos. Sin embargo,
una vez infectada su célula receptora, la célula infectada
procede a sintetizar la proteína del envoltorio del virus y el ADN
(o ARN) viral. Esto lleva a la muerte de la célula receptora y la
producción de muchas réplicas del virus infectante... de
allí que si bien es capaz de replicarse mediante la maquinaria celular
de otra célula, no se le suele considerar vivo.
Al no tener vida propia, los virus
pueden sobrevivir en latencia durante mucho tiempo, aun fuera de la célula
receptora. Su destrucción descansa sólo en la destrucción
molecular de su ADN (o ARN).
La importancia de los virus reside
tanto en el impacto que tienen sobre otras formas de vida (enfermedades)
como en la utilidad que ha tenido su forma de acción para la ingeniería
genética. No es del caso discutir aquí los aspectos prácticos
de la ingeniería genética, ya que por el momento las aplicaciones
industriales se limitan a la producción de drogas y reactivos finos
más que a tecnologías ambientales; sin embargo, es importante
destacar que la ruta a la modificación genética de bacterias
(típicamente Escherichia coli) para realizar la producción
de interés, se basa en mecanismos de modificación del gene
bacteriano que no son del todo distintas a la modificación viral.
Para el interés ambiental, existe
un virus que se suele utilizar como indicador de contaminación
fecal: es el Colifago somático y suele encontrarse relacionado con
las caracterizaciones de calidad microbiológica de aguas.
-
Algas
Exceptuando el alga azul-verde, que como
se mencionó más arriba es hoy en día considerada una
bacteria (en particular una cianobacteria), las algas tienen un
núcleo rodeado por una membrana (la membrana nuclear) y son, por
lo mismo, un protista eucariote (tienen núcleo diferenciado).
El tamaño de las algas recorre
desde las microscópicas algas del fitoplancton, unicelulares,
hasta las grandes algas marinas multicelulares. En todo caso, las
algas son más grandes que las bacterias, de estructuras mucho más
sofisticadas y altamente organizados. La mayoría de las algas son
fotosintéticas; sin embargo, tienen la capacidad de utilizar energía
química en ausencia de luz (durante la noche, consumen orgánicos
y oxígeno y generan CO2; es decir, todo lo contrario
de su metabolismo en condiciones iluminadas). Los pigmentos fotosintéticos
se encuentran en corpúsculos diferenciados llamados plastidios,
cloroplástos
o cromatóforos. Como se indicó antes, las algas son
fundamentales para la cadena alimenticia acuática; además,
producen la mayor parte del oxígeno atmosférico libre, como
un subproducto de la fotosíntesis necesaria para proveer su propia
vida.
Las algas se clasifican según
sus diferencias morfológicas, de tamaño y de composición
química, diferencias que -de paso- son pronunciadas. La tabla 2.5.1
despliega las clasificaciones típicas de las algas.
Tabla 2.5.1: Subdivisión
de las algas y su importancia
| Nombre usual |
Taxonómico |
Importancia |
| Algas Verdes |
Chlorofita |
Productor de
oxígeno libre; antecesor de plantas. |
| Euglenoides |
Euglenofita |
Sin paredes;
posible forma de transición evolutiva entre algas y protozoos. |
| Diátomas |
Cyclotella |
Productores
de oxígeno, vitamina A y D; tierra diatomacea útil para filtros
y abrasivos. |
| Dinoflagelados |
Pirrofita |
Envenenan mariscos |
| Algas rojas |
Rodofita |
Productora
de agar |
| Algas café |
faeofita |
Alga marina,
fuente de yodo, sosa; alimento; fertilizante; alginatos. |
Si bien las algas son los productores
primarios de alimento en la cadena alimenticia acuática, pueden
también producir problemas en los sistemas de agua potable ya que
contribuyen al olor y el sabor; pueden bloquear el paso de agua en tuberías;
bloquear medios filtrantes y demandar alta clorinación. El excesivo
desarrollo lleva al florecimiento de algas, que forma una capa (sábana)
de material orgánico cuyo aspecto estético es inaceptable
también para aguas recreacionales.
-
Hongos
Los hongos son prototistas no-fotosintéticos,
subdivididos en 3 grupos:
Mohos: hongos filamentados
que crecen desarrollando estructuras tipo filamento o hebra, llamados hyphae
y que terminan formando una masa llamada micelio (mycelium, mycelia).
El micelio vegetativo penetra en el sustrato y absorbe nutrientes mientras
que el micelio reproductivo es capaz de formar estructuras reproductivas,
como p.e. las esporas. Los mohos tienen aplicaciones importantísimas
en la industria farmacéutica, por ejemplo, en la producción
de penicilinas.
Levaduras: hongos no
filamentados, unicelulares, de tamaño mayor que las bacterias (5
a 30 µm de largo y de 1 a 5 µm de ancho), de formas ovoide,
ellipsiode o esférica. Su distribución en la tierra es de
amplia cobertura. Su reproducción puede o no ser sexuada; la reproducción
asexuada es por fisión binaria mientras que la sexuada es por formación
de esporas. Las levaduras son facultativas (crecen con o sin oxígeno),
a diferencia de los mohos. Las levaduras son muy conocidas para la humanidad;
se las utiliza en la producción de cervezas, pan; para la síntesis
de vitaminas, grasas y proteínas a partir de azúcares y nitrógeno
(amoníaco). Otras levaduras, como p.e. la Candida, causan
enfermedades graves.
Callampas: La forma más
tradicional del término "hongo" se refiere a la seta, conocida
en Chile como callampa. Es una forma altamente diferenciada y especializada
de hongo. El micelio está en el suelo, bajo la tierra, mientras
que la basidia se forma por sobre la tierra, en la estructura que típicamente
llamamos "el hongo".
La tabla 2.6.1 contiene algunos representantes
de estas especies de hongos. Se debe advertir que el listado podría
llegar a tener miles de especies, de modo que esta tabla no es más
que un ejemplo que viene al caso.
Tabla 2.6.1: Clasificación
simplificada de hongos
| Tipo |
División |
Comentarios |
| Mohos (filamentados) |
Phycomycetes |
Espora sexuada
o asexuada:
Mucor, Rhizopus |
| |
Fungi Imperfecti |
Sin etapa Sexuada:
Penicillium,
Aspergillus |
| Levaduras (no
filamentados) |
Ascomycetes |
Espora sexual
en sacos: Neurospora, Candida |
| Callampas (macroscópicas) |
Basidiomycetes |
Etapa sexual
en la basidia: Hongo común |
-
Protozoos
Los protozoos son las formas más altamente especializadas
de todas las formas unicelulares de vida. La gran mayoría
son no-fotosintéticos, se reproducen asexuadamente por fisión
binaria y no tienen pared celular. La mayoría de ellos son motiles
y -de hecho- su clasificación se basa en los mecanismos de locomoción.
Tabla 2.7.1: Los cuatro grupos de Protozoos y algunos
ejemplos clínicos
| Grupo |
Características |
Enfermedades
típicas |
| Mastigofora |
Se mueve por
flagella. Reproducción asexual (fisión binaria). |
Giardiasis;
Tricomonasis; Tripanosomiasis; Leismaniasis. |
| Ciliofora |
Se mueve y
se alimenta por cilios. Intercambio genético por conjugación.
Reproducción asexual, binaria, tranversa. |
Balantidiasis. |
| Sarcodina |
Se mueve y
se alimenta por pseudópodos. Reproducción asexual, binaria. |
Disentería
amébica. |
| Sporozoa |
No es motil.
Parásito intracelular. Reproducción en ciclo sexual y asexual.
Suele requerir series de huéspedes para su ciclo vital. |
Malaria; Toxoplasmósis;
Pneumocistis (forma de pneumonia). |
Muchos protozoos, según se reseña
en la tabla 2.7.1 son conocidos desde hace mucho tiempo por las ciencias
de la medicina (donde la microbiología es una de estas ciencias).
Se puede esperar encontrar protozoos en prácticamente cualquier
hábitat húmedo, y su alimentación suele ser saprofítica
(consume substratos disueltos) y se alimenta de bacterias; sus tamaños
son variados: desde unos pocos micrones hasta varios cientos de micrones.
En condiciones adversas, se encapsulan en un quiste de duras paredes y
logran sobrevivir aun en condiciones extremas.
El género sarcodina,
p.e., tiene una membrana celular continuamente cambiante debido a la generación
de pseudópodos (extremidades falsas); este mecanismo, típico
de la amoeba, le sirve para el desplazamiento y para englobar su
alimento e ingresarlo al citoplasma. Estos mismos pueden ser patógenos
a los humanos, p.e. la Entamoeba histolica es un patógeno
muy difundido que causa la disentería amébica. Algunos protozoos
son fotosintéticos, p.e. Euglena, y comparten algunas características
con las algas (como, por ejemplo, producir dificultades a la producción
de agua potable). Otros, son parásitos, p.e., la Trypanosoma
es un parásito de la sangre y produce somnolencia en humanos. Los
protozoos parasitarios pueden ser motiles (suctoria) o no motiles (sporozoa).
Entre los no motiles destacan cuatro especies de Plasmodium, causantes
de la malaria y que viajan mediante un mosquito (el mosquito Anopheles
hembra).
Aplicaciones
de la Microbiología a Ciencias Ambientales
Microbiología
del suelo (Tierra)
La tierra,
(149,600,000 km (1.00 AU) del Sol, diametro: 12,756.3 km, masa:
5.9736e24 kg), como es sabido, está formada por diversas
capas, entre las que se puede distinguir con bastante claridad,
la capa más externa como aquella que soporta, junto con los océanos,
las diversas formas de vida. En particular, llamaremos aquí "suelo"
a la capa más externa, aquella capaz de soportar el crecimiento
biológico; esta capa es una fracción
ínfima del radio de la tierra y, con pocas excepciones, no
ocupa más que unos pocos metros (ver diagrama en la figura
3.1.1).
El suelo actúa como sistema de reciclaje
de materiales orgánicos ya que en él ocurren tanto
la vida (animales, plantas, protistas como pseudomonas)
como la recepción de los restos de ella una vez terminado un ciclo
vital y los desechos permanentes del metabolismo (esto es cierto, además,
para los seres voladores). Es en el suelo donde la actividad
microbiana transforma los desechos en los componentes del propio
suelo. Los diversos ciclos químicos (del carbono, del nitrógeno,
del azufre, etc.) se realizan mediante actividad microbiana y sin
ellos la vida en la tierra desaparecería después de
sintetizar un gran número de moléculas orgánicas de
variada diversidad.
La composición del suelo (figura
3.1.1) varía de un lugar a otro pero, en general, siempre se
podrán distinguir cinco fracciones principales:
-
Partículas inorgánicas
minerales: Principalmente, aluminio y silicio; en fracciones menores,
todos los demás minerales. Los tamaños varían desde
las partículas de cerámica (2 µm) hasta granos de arena
y piedras. La proporción de uno y otro material determina la capacidad
de retención de agua, la estructura, la permeabilidad para el aire
y la retención de nutrientes. El tipo más común de
suelo consiste, precisamente, de inorgánicos y lleva ese mismo nombre.
-
Residuos orgánicos:
Constituido por los residuos de plantas y animales, en diversos estados
de descomposición, que llegan a un estado estable, conocido como
el humus. Se llama "suelo orgánico" a aquel formado principalmente
por humus.
-
Agua: El agua es fundamental
para la actividad microbiana. La cantidad de agua presente en el suelo
depende de la pluviosidad, la estructura del suelo y la población
microbiana, entre otros. El agua puede ser retenida en los poros entre
partículas adyacentes (suelos saturados) o absorbida en la superficie
de las partículas (suelos secos). Los nutrientes se disuelven en
el agua y son así accesibles a los microorganismos.
-
Gases: Los tres gases de
mayor importancia para la vida en la tierra son el oxígeno, el nitrógeno
y el dióxido de carbono (naturalmente, existen muchos otros: p.e.
el ácido sulfhídrico y el metano). Los gases se alojan en
los espacios no ocupados por el agua y, en suelos saturados (inundados),
los gases se encuentran disueltos en fase líquida.
-
Sistemas biológicos:
Formado por los sistemas de raíces de plantas, los pequeños
animales (p.e. insectos), los microorganismos y sus sistemas comensales,
esta fracción del suelo constituye la vida en el suelo y produce
todas las transformaciones observables. En términos cuantitativos,
un gramo de suelo agrícola suele contener ¡2.500
millones de bacterias, 500 mil hongos, 50 mil algas y unos 30 mil protozoos!.
Se ha mencionado que, en términos cuantitativos,
las bacterias y los hongos forman la mayor parte de los microbios encontrados
en ambientes naturales, particularmente, en el suelo.
Dependiendo de las condiciones específicas del
suelo que se analice, se encontrarán bacterias
anaerobias o aerobias, heterótrofas o autotróficas
junto a hongos descomponiendo estructuras
poliméricas como la celulosa y la lignina de la estructura de las
plantas, todos ellos formando un concierto sorprendentemente bien adaptado
a las condiciones específicas de cada suelo. A lo largo de la evolución
del proceso de descomposición de sustratos, irán variando
las fracciones dominantes de unas u otras bacterias u hongos, en una cooperación
cuya consecuencia final es la mantención de los ciclos vitales.
Como la mantención de una forma de vida particular
(p.e. una especie de bacteria) depende de la disponibilidad de todos (y
cada uno de) sus requerimientos vitales (ver tabla
2.2.2 p.e.) y de la ausencia de los elementos que le resultan nocivos
(inhibitorios), se encontrará que en un hábitat particular
del suelo existirán aquellas especies mejor adaptadas a las condiciones.
Las condiciones de mayor incidencia son: humedad
disponible para la solubilización de los nutrientes y desechos;
cantidad y calidad adecuada de nutrientes y
condiciones
físico químicas adecuadas (se suele hablar del pH
y la temperatura, principalmente, pero las condiciones de Eh pueden ser
también importantes).
En cuanto a las aplicaciones
industriales más típicas que utilizan las propiedades
biológicas de los suelos (más allá de lo agrícola)
se debe tener en cuenta la compostación
y los rellenos
sanitarios. El típico relleno sanitario, reseñado en
corte en la figura
3.1.2, puede generar problemas ambientales si no es manejado con cautela
(olores y contaminación de aguas, de la napa por percolación
o superficiales por flujo superficial) y, por ello, se han establecido
cuidadosas normas de diseño y construcción que se pueden
encontrar en manuales, normas o especificaciones de diversos países.
Tomando el relleno sanitario
como un ejemplo de la diversidad de interacciones que ocurren en el suelo,
se puede destacar que no existe
un único microorganismo que pueda degradar
una gran diversidad de sólidos orgánicos húmedos (como
la basura domiciliaria) hasta metano y CO2 (ver figura
3.1.2). La operación la realizan, en realidad, un gran consorcio
de micro- organismos que realizan sus distintas actividades metabólicas
con el sólo fin de su propia vida; ocurre, eso sí, que los
productos
finales de la actividad de algunos microorganismos, descartados
a la fase líquida exterior a la célula, pueden ser los alimentos
de otros, enlazándose así en una compleja cadena que lleva
hasta los productos gaseosos observados. En una muy breve síntesis,
según se sintetiza en la figura
3.1.3:
-
en primer lugar, los orgánicos
insolubles (lípidos, polisacáridos, proteínas,
ácidos nucléicos, etc.) son degradados a especies orgánicas
solubles
por la acción de enzimas extracelulares, produciendo especies de
menor peso molecular que pueden cruzar la membrana celular y ser digeridos
por los microorganismos (ácidos grasos, monosacáridos, amino
ácidos, purinas, pirimidinas, aromáticos, etc.). A continuación,
-
las bacterias productoras de ácido
consumen estos productos solubles generando, entre otros productos y aparte
de las mismas células, ácidos volátiles (principalmente,
ácido acético [70%]; ácido propiónico; ácido
butírico y otros en menor cuantía). Sólo una vez producidos
los ácidos volátiles
-
las bacterias productoras de
metano pueden realizar su metabolismo que lleva estos ácidos
volátiles hasta metano (CH4) y dióxido de carbono
(CO2), o CO2 y H2a CH4
como el Methanococcus
jannischii además de generar, naturalmente, células.
Es importante destacar, además, que las primeras
etapas del proceso son aerobias, mientras que las últimas,
particularmente la generación de metano, ocurre sólo en condiciones
estrictamente
anaerobias;
es decir, no sólo concurren muy diversos microorganismos en el proceso
sino que, además, las condiciones físico
químicas varían a lo largo del proceso.
Aquellas industrias cuya descarga es un sólido
de características orgánicas pueden recurrir a la
metanogénesis (en alguna operación similar al relleno sanitario
o, más usualmente, en reactores diseñados específicamente);
a la compostación (tanto abierta al ambiente como en reactores),
a la hidrólisis de celulosa; y/o a algunas otras alternativas más
complejas. En todo caso, los descartes sólidos
pueden ser tratados biológicamente sólo en presencia de
agua.
-
Microbiología de Líquidos
en la biosfera
En ausencia de aguas industriales y urbanas (es decir,
sin considerar las actividades netamente antropogénicas de canalización
de aguas) todas las aguas provienen de precipitaciones, sea en forma de
lluvia o de hielos fundidos.
Durante los primeros minutos de precipitación,
la lluvia disuelve y arrastra los gases, las partículas y los microorganismos
presentes en el aire. Posterior a ello, la lluvia consiste de agua pura.
Las aguas llegan al suelo, donde son absorbidas por tejidos vegetales y
por las partículas del suelo. La fracción no absorbida puede
fluir en la superficie o percolar hacia las napas subterráneas;
en cualquier forma, su destino final serán ríos, lagos, lagunas
y -finalmente- el océano.
Las aguas de la napa subterránea
son relativamente asépticas (tienen
escasos microorganismos suspendidos), ya que el agua se filtra al percolar
desde el suelo y, más aun, no está expuesta a la luz. Naturalmente,
si la napa contacta áreas ricas en minerales de hierro, puede que
se encuentren acidófilos oxidantes que la contaminen. Por otra parte,
puede ocurrir que la napa reciba contribuciones de agua superficial no
percolada, a través de grietas u otro tipo de canalizaciones y,
en este caso, la napa se contaminará con microorganismos típicos
de la superficie.
Las aguas superficiales,
por otra parte, se van cargando de muy diversos microbios a lo largo de
su viaje por tierras agrícolas. No sólo eso, además
se pueden cargar de nutrientes, nitritos, fosfatos, etc. Por todo ello,
en ausencia de descargas tóxicas, las aguas
superficiales suelen tener una alta actividad microbiana. La
actividad biológica en las aguas, p.e. de una laguna, establece
cadenas biológicas similares a las descritas para el suelo, donde
los productos de unos es el sustrato
de otros, además de producirse relaciones
predador-presa; con fines ilustrativos, según se describe
en la figura
3.2.1, en una laguna de estabilización
de aguas servidas, la carga orgánica presente en las
aguas servidas que entran es "tratada" por un consorcio biológico
donde participan diversos microorganismos, en una visión simplificada:
-
las bacterias (aerobias o
anaerobias) degradan la carga orgánica produciendo una cantidad
notable de nuevas bacterias (ya que la carga orgánica es alta y
suficientemente biodegradable) y una alta concentración de CO2.
Si las bacterias son aerobias, requerirán una gran cantidad de oxígeno
(precisamente aquel determinado por la
demanda
bioquímica de oxígeno, usualmente abreviado a
DBO
y medido en cinco días DBO5)
para su respiración.
-
Las algas, por otra parte,
pueden crecer en la superficie de la laguna, allí donde la intensidad
de la radiación solar es alta. Al crecer en condiciones iluminadas,
consumen CO2 (el producto de descomposición
de la carga por las bacterias) y generan oxígeno
(requerido por las bacterias
para su respiración aerobia). Así, las
algas y las bacterias interactúan permanentemente mediante
estos dos gases y el producto neto será la disminución de
los orgánicos disueltos y la obtención de más algas
y bacterias. Luego,
-
los protozoos predan (se
comen) a las bacterias y a las algas, predación que arroja como
resultado neto un cierto equilibrio poblacional.
Los mecanismos de regulación
de cuantas bacterias, algas y protozoos, de diversas especies, existan
es de la mayor trascendencia; pero la observación empírica
es que se mantiene un cierto equilibrio que arroja, finalmente, un efluente
(salida) más
limpio de carga orgánica y de
microorganismos de origen fecal que el afluente original. Que
el efluente cumpla los requisitos de diseño suele ser más
un problema de manejo de la fluidodinámica y del tiempo de residencia
que un problema de equilibrio de poblaciones microbianas.
Desde un punto de vista de diseño,
la participación de distintos microorganismos en el proceso podría
ser modelado mediante sistemas de comensalismo,
competencia
por sustrato, relaciones predador-presa, y
una cantidad de otras formas de interrelación.
El sistema resultante sería complejo, sin duda.
Con un propósito meramente ilustrativo, considérense
la relación existente entre bacterias, protozoos y carga orgánica.
Supongamos, p.e., un sólo tipo de bacteria (concentración
X
bacterias/L)
que se supondrá creciendo en función de un único sustrato
(es decir, se supondrá que los otros sustratos están en exceso,
concentración S g/L) que entra a tasa
constante (p.e. el carbono total del afluente a flujo
F
L/s). Supóngase que las bacterias sean predadas por un sólo
tipo de protozoo (concentración
P protozoos/L)
que las engulle en función de la concentración de las propias
bacterias.
Se pueden suponer formas matemáticas razonables
para el muy simple sistema propuesto y se verá que el sistema matemático
resultante presenta -para ciertos valores de los parámetros que
definen el modelo- soluciones oscilatorias e incluso, inestables.
La oscilación podría ocurrir,
por ejemplo, si los protozoos aumentan en número debido a una acumulación
de bacterias; el aumento en P producirá,
a la larga, una escasez de bacterias y por lo tanto de protozoos; el decremento
en X, a su vez, producirá una elevada
concentración de sustrato, incentivando así el aumento de
X;
luego X crece nuevamente mientras los protozoos
estén desapareciendo por falta de su comida X.
Etcétera...
Las aguas, sean ellas superficiales o subterráneas,
de acuerdo a la ley, se deben caracterizar
en su calidad previo a su posible utilización.
La norma chilena NCh409, por ejemplo, especifica las características
del agua potable mientras que la norma NCh1333 define las características
que debe cumplir un caudal para ser utilizado en riego, recreación,
estética, bebida de animales, etc. (es decir, todo otro uso, más
allá de agua potable).
En cuanto a la calidad microbiológica
del agua, esta se suele caracterizar sólo contando los coliformes
fecales y, a veces, los virus colifago;
ambos microbios son de origen humano (pertenecen a la flora intestinal)
y permiten determinar si un caudal ha tenido contacto con aguas servidas.
Sin embargo, existen las técnicas para medir multitud de otros microorganismos
y de hecho los hay muy diversos en el agua. El objetivo de determinar la
presencia (y cuantía) de los microorganismos indicadores
de contaminación fecal busca, en realidad, determinar que un caudal
particular no contenga microorganismos causantes
de enfermedades entéricas (digestivas) infecciosas. El espíritu
de las normas mencionadas es la protección de la salud de la población
que utiliza las aguas.
Las enfermedades transmitidas por las aguas son diversas.
El mecanismo de transmisión es por ingestión de patógenos
acarreados por el agua. Algunas de las enfermedades de mayor incidencia,
cuya transmisión es desde las feces de un enfermo al sistema digestivo
del nuevo infectado son:
-
las gastroenteritis en general (bacterianas, e.g. E.
coli),
-
la gastroenteritis viral,
-
la giardiasis,
-
la shigelosis,
-
la hepatitis A,
-
la campilobacteriosis,
-
la salmonelosis,
-
la tifoidea (Salmonella
typhimurium),
-
el cólera (Vibrio
cholerae),
-
la amebiasis,
-
y muchas más.
Cada una de estas enfermedades
es causada por un microorganismo específico que podría
estar presente en el agua si algún
enfermo hubiese depositado sus feces en el caudal. El punto de interés
aquí es que
| si el enfermo descargó
el microorganismo patógeno de su enfermedad en el cauce, entonces,
también descargó los microorganismos de la flora normal del
intestino. |
De allí que en lugar de intentar medir la presencia
y cuantía de cada posible patógeno presente en un agua es
más simple medir los indicadores de
contaminación fecal. Los indicadores estarán presentes en
el caudal de agua contaminada con feces, independientemente
de si existen o no patógenos.
Un indicador debiera cumplir con ciertos requisitos
"ideales". Un buen indicador sería aquel:
-
cuya ausencia implique la ausencia de patógenos
entéricos;
-
cuya densidad se relaciones con la probabilidad de presencia
de patógenos;
-
cuya sobrevivencia fuese ligeramente superior a la de
los patógenos.
Tal como suele ocurrir con los ideales, tal indicador
no existe (al menos por ahora). En Chile se suele utilizar sólo
los coliformes fecales y coliformes
totales, con alguna tendencia a medir la presencia del virus
colifago.
En otros países se ha reglamentado medir, además, la presencia
del Clostridium perfringens.
En cuanto a la sobrevivencia de los patógenos
en el agua, esta no es distinta a la sobrevivencia de otros microorganismos
en el agua. Los mecanismos que "matan" bacterias que no formen esporas,
pueden ser naturales o acelerados. Cualquiera sea el proceso de "desactivación
celular", este se basará en fenómenos que:
-
rompen células,
-
denaturan proteínas,
-
envenenan células o
-
bloquean el ciclo vital.
Por ejemplo, la remoción de patógenos en
las etapas finales de tratamientos de aguas servidas se suele realizar
con operaciones que producen la ruptura de la membrana celular por una
fuerte oxidación química, por
clorinación
o por ozonización. Similarmente,
se puede recurrir a la radiación UV
del caudal, que destruye las células por ruptura química
exhaustiva de su material cromosomal. Las lagunas de estabilización,
en cambio, remueven microorganismos patogénico por ausencia
de sustrato (comida) y por exposición
a microorganismos que los pueden engullir.
El diseño mismo de las lagunas
de estabilización descansa sobre la cantidad de tiempoque
debe ser retenida un agua contaminada para
que se observe la reducción en la cuantía de indicadores
en una magnitud determinada.
Por ejemplo, una entrada de aguas servidas típica
tendría unos 107 coliformes fecales
(CF) en 100 mL del caudal (curiosamente, se ha sostenido en el tiempo el
hábito de expresar la densidad celular de estas bacterias con relación
a un volumen de 100 mL y no por mL ni por
litro). El requerimiento de diseño podría ser la obtención
de un caudal reutilizable en riego, es decir que no tenga más de
103
CF/100 mL, según especifica la norma NCh1333. Se requiere,
por tanto, reducir los CF en cuatro ordenes de magnitud (¿cuanto
se reducirá el cólera?, ¿la
Salmonella typhi?,
¿la
Shiigella?). Es decir, en términos muy sintéticos,
se diseñará para llevar un número de microorganismos
del afluente, N0, a las especificaciones
de la ley, NE, en el efluente.
La experiencia en sistemas estáticos (no continuos, batch) indica
que el decaimiento de los CF puede ser más o menos bien representado
por una ley de primer orden:
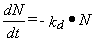
Luego, según el tipo de flujo que se produzca
en la laguna en cuestión (nota: las lagunas son un sistema continuo
donde entra y sale líquido en todo momento), se tendrá una
de las tres tasas de decaimiento (se presentan los modelos correspondientes
a flujo pistón y mezcla completa; existe también
el modelo para flujo disperso):

donde N0 es
el número de coliformes fecales de entrada
y NE es el número de
coliformes
fecales de salida (efluente);
kd
es la constante de decaimiento de primer orden
y t
(letra griega tau) es el tiempo de residencia
del líquido en el sistema (volumen dividido por flujo).
La constante de decaimiento, como se ha descrito, se
puede medir en un sistema batch. Diversas investigaciones ha llevado a
establecer una relación clara entre la tasa de decaimiento de los
CF y la temperatura. Bastará medir la constante a 20 ºC y a
otra temperatura (típicamente 15 ºC) para encontrar un parámetro
de ajuste, q ,
y establecer una ley exponencial simple: kd = k20×q(T-20),
con T en grados centígrados. Con todo lo anterior, se puede, ciertamente,
diseñar cualquier aplicación de remoción de bacterias
si
se conoce su constante de decaimiento.
Como dato de referencia, el sistema experimental de
lagunas
Esmeralda (EMOS S.A.), que constaba de dos lagunas en paralelo seguidas
por una tercera en serie con las dos primeras, trata las aguas servidas
de unos 12.200 habitantes, a un caudal medio de 22,6 L/s. Ocupa un área
de unos 60 mil m2 de lagunas a
una profundidad de 1,8 m. Los tiempos de residencia diseñados eran
de 19,1 y 18 días para las dos lagunas
primarias y de 11,4 días para la secundaria.
El estudiante podrá calcular cuantos ordenes de magnitud se reducirán
las densidades de microorganismos indicadores en este sistema... :-)
Desde el punto de vista del tratamiento de las aguas
servidas, que son colectadas en el alcantarillado, las descargas
industriales que se viertan al alcantarillado público no
deberían contener especies que pudiesen resultar nocivas
para el sistema microbiano que se establece en las plantas de tratamiento.
Es decir, una descarga industrial a alcantarillado no debiera interferir
con los procesos aguas abajo. Tampoco serían aceptables las descargas
de especies que impidan la reutilización de las aguas por causa
de elementos que, cruzando incólumes las plantas de tratamiento,
impidan su uso, p.e., para riego. Por otra parte, tampoco sería
aceptable descargar elementos que precipiten en los ductos de aguas servidas
o que los corroan.
Desde un punto de vista biológico, es importante
contestar el cómo saber si una descarga:
-
resultará nociva para los
tratamientos aguas abajo;
-
qué elementos pueden cruzar
las plantas de tratamiento y ser luego nocivos para el suelo o para quienes
consuman los vegetales regados con esas aguas;
-
qué indicadores de contaminación
industrial se pueden utilizar;
-
qué límites se deben
fijar para la descarga de cuales elementos.
Son todas preguntas que la sociedad ha contestado con
diversos argumentos, diversas orientaciones y, sobre todo, muy diversos
métodos técnicos y plataformas políticas.
-
Descargas Industriales
Se discutió más arriba (ver figura
1.1.1) que cerrar explícita y totalmente el balance de cada
especie que participa en un proceso resulta difícil y, probablemente,
inútil. La práctica dicta más bien la necesidad de
utilizar ecuaciones de balance para las especies
que entran directamente en el proceso y balancear sólo en
forma global el resto de las especies. Estas últimas aparecerán
en los efluentes y deberán ser tratadas en procesos suficientemente
flexibles como para compensar el desconocimiento de las ecuaciones analíticas
de balance. Naturalmente, se supone aquí que el diseñador
a realizado todos los esfuerzos para minimizar la contaminación.
Se ha reseñado que los materiales descartados
de cualquier proceso pueden ocurrir en diversas fases. Sin embargo, la
fenomenología natural es tal que -finalmente, luego de muy diversas
interacciones- los descartes llegarán a la
fase líquida (por medio de lluvias, arrastre, solubilización,
etc.). Así como muchos materiales disueltos en agua pueden resultar
útiles para el desarrollo de microorganismos, otros elementos pueden
resultar definitivamente nocivos; sin duda que una cantidad importante
de especies disueltas que resultan nocivas para algunos microorganismos
pueden resultar benéficas para otros, en este caso se observa más
bien un desequilibrio en las especies microbianas
dominantes, llegando incluso a determinar los tipos de peces que pueden
estar presentes.
Además, tanto los nutrientes
en exceso como los inhibidores pueden resultar nocivos para las
especies microbianas. La excesiva descarga de nutrientes (nitrógeno,
fosfato, sales, etc.) tanto como la descarga de compuestos orgánicos
promueve un exceso de actividad celular, a
un punto tal que todo el oxígeno atmosférico que se disuelve
en el líquido es rápidamente consumido por los microorganismos
que habitan el primer par de centímetros
de la superficie del agua, produciendo condiciones
anóxicas, con el consecuente desarrollo de microorganismos
anaerobios; los anaerobios son molestos pues tienden a producir compuestos
malolientes. La descarga de inhibidores de las poblaciones microbianas
típicas de un cauce sano imposibilitan su desarrollo a la vez que
podrían promover el desarrollo de especies no deseables.
La figura
3.3.1 sintetiza, en términos muy globales, como el alto consumo
de oxígeno genera condiciones anóxicas en un río que
recibe descargas orgánicas y/o de nutrientes:
-
el oxígeno atmosférico
se disuelve en el líquido (oxígeno disuelto) a una tasa regulada
por las propiedades de transporte del líquido;
-
el oxígeno disuelto es utilizado
por los microorganismos aerobios para metabolizar los orgánicos
y los nutrientes y generar más células;
-
el número de células
llega a ser tan alto que el oxígeno no alcanza a penetrar más
que unos pocos centímetros; así,
-
el oxígeno se hace deficitario
y se pueden desarrollar los anaerobios;
-
producto de la digestión
(aerobia y anaerobia) a lo largo del cauce, la carga excesiva es consumida
y se logra restablecer el nivel de oxígeno disuelto original.
Claramente, más que legislar respecto
de la producción de regiones anóxicas en las aguas superficiales,
convendría regular la máxima concentración
de orgánicos consumidores de oxígeno por vías
microbiológicas; tal definición podría resultar
en una variable cuantificable por métodos simples ya que, simplemente,
se trata de medir el potencial de consumo de oxígeno
de una descarga determinada.
-
La Demanda Bioquímica
de Oxígeno: expresión de la actividad microbiana
En particular, la demanda
bioquímica de oxígeno (DBO)
especifica la cantidad de oxígeno (en
miligramos de oxígeno por unidad de volumen, usualmente mg/L) que
sería necesario para degradar, por
oxidación a dióxido de carbono, la carga orgánica
de una muestra, mediante la acción de microorganismos.
Es de interés comprender como se determina la DBO, en un test que
toma cinco días (de allí el término DBO5,
que fija los límites legales máximos a las descargas de orgánicos).
-
En una
jarra de vidrio (la botella de DBO, puede ser vista también
como un reactor estático o batch) se vierten 300 mL de muestra con
algún grado de dilución (se usan varias diluciones simultáneamente),
saturada en oxígeno y con un cultivo de microorganismos variados,
capaces de digerir aeróbicamente la carga orgánica (en muchos
casos se agrega un cultivo estándar).
-
La jarra
se incuba a 20 ºC durante
5 días, al final de los cuales
se mide el oxígeno remanente. Por diferencia (es decir, la concentración
de saturación de oxígeno que existía al inicio del
análisis menos la concentración remanente a los 5 días)
se determina el oxígeno consumido por los microorganismos para degradar
materia orgánica de la muestra. Naturalmente, no toda la carga orgánica
habrá sido degradada. Además, se debe corregir por el factor
de dilución aplicado.
|
A partir de la DBO5 se estima
la DBO terminal (DBOu, por "últimate"), la que será,
generalmente, un 30 a 40% mayor que la DBO5. Este valor determina
el requerimiento de oxígeno que presenta un
caudal particular. Si la magnitud de la DBO es excesiva producira
condiciones anóxicas en el cauce receptor.
Además, esta cantidad de oxígeno
es, precisamente, la que debiera ser entregada
al sistema líquido en el plazo en que la carga debe ser digerida.
Este es el punto en que una carga orgánica puede ser excesiva. Sea,
por ejemplo, el caso de un río. El transporte de oxígeno
desde la atmósfera será determinado por las propiedades de
transporte desde la atmósfera (con una presión parcial dada
de oxígeno, típicamente 20%) a la fase líquida; usando
una de las teorías más simples de transporte, la tasa de
transporte de oxígeno desde la atmósfera al líquido
será proporcional a la fuerza motriz dada por la diferencia entre
la concentración máxima posible de oxígeno en el líquido
(la concentración de saturación, C*) y la concentración
actual (C); la constante de proporcionalidad está determinada por
el área de contacto entre gas y líquido (a) y una constante
de transporte que denominaremos kl:
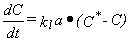
La población microbiana, a su
vez, consume oxígeno a una tasa que es proporcional al número
de células presentes (suponiendo una sola cepa, se hablaría
de la tasa de respiración, r).
La población es mixta para el caso de un río, pero como elemento
conceptual, se puede considerar una variable X que representa las células
aerobias y una tasa de respiración r, por unidad de masa de X. Así,
el consumo de oxígeno disuelto en la fase líquida estará
dado por:

La población, X, a su vez estará
regida por ecuaciones de crecimiento que dependen del sustrato (S)
que consumen, mediante ecuaciones del tipo Monod, donde la tasa de crecimiento
(aumento de microorganismos) está dada por una parábola que
depende de una velocidad máxima de crecimiento (µmax)
y por una constante de afinidad con el sustrato (Ks):
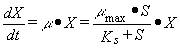
Naturalmente, la ecuación de
crecimiento de Monod recién descrita
rige cuando S es el sustrato limitante.
Si el oxígeno es deficitario, será necesario escribir la
ecuación con S representando al oxígeno disuelto y no al
carbono orgánico u otro nutriente. De hecho, si la DBO5
supera los límites establecidos, la limitación al crecimiento
será por oxígeno ya que la kla será demasiado
pequeña para la tasa de respiración que se requeriría
(de allí que se deben diseñar sistemas de aireación
forzada, incluso con oxígeno puro en algunos casos).
-
Detalles matemáticos
de modelamiento de especies
Crecimiento
Exponencial
Según se indicó en la
sección de microbiología básica, en general, se observa
que si hay crecimiento, la cantidad de organismos que se desarrolla en
una unidad de tiempo es una función de la cantidad de los mismos
organismos presentes al tiempo inicial: C = f(N),
donde C es la tasa de crecimiento y
N
la cantidad de organismos. Para la función
f,
se deberán realizar observaciones experimentales directas sobre
la población de interés; durante muchos años de investigación
se han llegado a establecer algunas funcionalidades típicas.
El caso más simple, válido
solo en rangos pequeños de interpolación, se da cuando la
tasa de crecimiento (reproducción) es directamente proporcional
a la cantidad de microorganismos (N)
presentes:
dN / dt =
C = k N
donde k es la constante de proporcionalidad.
La integración de esta ecuación lleva a una exponencial:
N=N0×exp(k·t),
donde N0 es la población a tiempo cero y t
es el tiempo. Naturalmente, este modelo resulta de una simplicidad extraordinaria
puesto que no existe el caso de poblaciones que crezcan
eternamente hasta un número infinitamente alto de elementos
(como ocurre con la función exponencial). Más allá
de la constatación experimental, el caso es que los microorganismos
pueden morir, al igual que pueden crecer, según la disponibilidad
de alimentos. Se debe, por lo tanto, introducir un mecanismo de desactivación
o muerte de células.
Decaimiento
exponencial:
Nuevamente, el modelo más simple
supone que la tasa de decaimiento es directamente proporcional al número
de organismos presentes. En este caso, la ecuación resultante es
similar a la anterior pero lleva un signo menos delante de la constante
de decaimiento
kd: N=N0×exp(-kd·t).
Crecimiento
inhibido (logístico):
La observación experimental
indica que el número de organismos no crece indefinidamente
sino que se estaciona en algún valor límite. Si el valor
límite (L) es conocido, entonces es posible modelar el crecimiento
mediante:
dN / dt =
k·N·(L - N)
Esta ecuación puede ser integrada
analíticamente:
N = (N0·L)/(
N0+ {L-N0}×
exp[-L·K·y] )
Esta ecuación se utiliza a menudo
en modelamiento del crecimiento en condiciones realistas de limitaciones
de sustrato (alimentación, nutrientes, etc.) y para la propagación
de infecciones.
El modelo logístico se suele
expresar mediante una constante de crecimiento que contempla tanto el crecimiento
como la mortalidad, sintetizándolas en una sola constante (r);
similarmente, más que buscar un valor para el límite de crecimiento
(L), se suele definir una capacidad del medio de cultivo
(es decir, cuanto organismos podrían desarrollarse a partir de las
condiciones alimenticias que se han puesto en el medio de cultivo), capacidad
representada por K. Entonces:
dN / dt =
r·N·(1 - N/K)
podrá deducirse que r
= k×
L en
concordancia con la ecuación logística original.
Este modelo es también útil
para el modelamiento de especies animales, excepto que la capacidad ambiental
para el desarrollo de herbívoros (K) no es constante a lo
largo de un año. En este caso se suele utilizar una capacidad variable.
Puede ser de interés mencionar, por ejemplo, la capacidad anual
de un ecosistema, modelada mediante
K = k×
seno(2×p×
t/tl ) + K0
donde K es la capacidad del
medio, k una amplitud, tlel
período del ciclo (si es estacional, 365 días, 52 semanas,
etc.) y K0 la capacidad promedio.
Modelos de
competencia entre especies:
El modelo de Gause y Witt, basado en
los modelos de Lotka y Volterra, plantea el crecimiento de dos especies
simultáneamente. Sin definir exactamente el efecto de una especie
sobre la otra, es posible postular "factores" de impacto de una especie
sobre la otras. El modelo se basa en ecuaciones logísticas como
las recién analizadas:
dN1
/ dt = r1×N1
- r1×N12
/ K1 - a×
r1×N1×N2
/ K1
dN2
/ dt = r2×N2
- r2×N22
/ K2 - b×
r2×N1×N2
/ K2
donde N1 y N2
son las densidades de población (número de organismos por
unidad de área o volumen) de las especies 1 y 2; r1
y r2 las tasas de crecimiento de cada una; K1
y K2 las capacidades de soporte del ambiente para
cada especies en ausencia de la otra;
a
es una constante que relaciona el efecto que tiene el crecimiento de la
especie 2 sobre el crecimiento de la especie 1 y b
es una constante que relaciona el efecto que tiene el crecimiento de la
especie 1 sobre el de la especie 2.
En lenguaje menos matemático,
las ecuaciones expresan que la tasa de crecimiento de una especie está
dada por su crecimiento ilimitado, menos la limitante ambiental
propia y menos el efecto de la interacción de las dos especies.
Relaciones
predador-presa:
Lotka propuso un modelo de la interacción
predador (el que come)- presa (el comido) para poblaciones homogéneas
en el año 1923. Curiosamente, Volterra propuso el mismo modelo en
forma independiente en el año 1931... desde entonces, el modelo
se conoce como Lotka y Volterra.
Describiendo la densidad de población
del predador por P y la densidad de las presas por N, el
modelo postula:
dN / dt =
r× N - rp×N×
P
dP / dt =
a×
rp×N×
P - kd×P
donde r es la constante de crecimiento
de las presas; rp es la constante de predación
(predadores producidos por presas capturadas) y kd es
la tasa de mortalidad de los predadores.
Un conjunto típico de valores
para este modelo es:
N0=
1500; P0= 50; r= 0,1; rp=0,002; kd= 0,2;
a
= 0,1.
Estos valores pueden ser utilizados en
un simulador simple, por ejemplo mediante el programa BASIC que se incluye
en la figura 3.5.1, y arrojará los resultados
incluidos en el gráfico de la figura
3.5.2. A pesar de la simplicidad del modelo, es evidente que existen
oscilaciones con tan sólo considerar dos especies. Si se intentara
modelar un ecosistema la posibilidad inestabilidades
(oscilaciones) sería aún mayor. Es del mayor interés
evaluar, con este simple modelo, cuales son las condiciones (metemáticas)
para que el sistema fuese estable; para que el sistema oscile establemente;
para que la oscilación del sistema fuese amortiguada o infinita;
y, sobre todo, bajo que condiciones el sistema finalmente
decae y desaparecen las presas y/o los predadores.
Advertencia:
Todos los modelos tienen, sin duda,
dificultades para representar los datos experimentales. El problema es
que los rangos de validez de las hipótesis utilizadas para construir
las ecuaciones son extrictos y no pueden ser excedidos. Es un asunto de
verificación experimental práctica que modelo puede ser útil
para cada situación.
Por otra parte, las situaciones realmente
interesantes desde un punto de vista de aplicaciones a efluentes líquidos,
no corresponden ni a cultivos de cepa pura (monocultivo, como los descritos
en la sección 2) ni a competencia o relaciones entre sólo
dos especies, como las descritas aquí.
En realidad, en un proceso cualquiera,
abierto al ambiente, participan muchas especies (al menos decenas, aún
para procesos muy restrictivos y de medios de cultivo altamente agresivos,
por ejemplo la lixiviación bacteriana a pH del orden 1,5).
A pesar de lo restrictivo y restringido
de los modelos listados más arriba, estos forman la base de la mayor
parte de los modelos biológicos en uso. La sofisticación
crece con el grado de precisión exigido por el modelo y no es extraño
llegar a los cientos de ecuaciones diferenciales. En la práctica
de diseño, en cambio, se opera con modelos donde las tasas son de
carácter global y los parámetros se ajustan empíricamente
en operaciones de laboratorio o piloto.
| Programa de simulacion Lotka Volterra
DEF fna (n,p) = r * n - rp * n * p
DEF fnb (n,p) = alfa * rp * n * p - kd * p
alfa = .1
kd = .2
rp = .002
r = .1
p0 = 50
n0 = 1500
paso = .01
OPEN "lotka.dat" FOR OUTPUT AS #1
FOR t = 1 TO 160 STEP 2
p = p0
n = n0
FOR ti = t - 1 TO t STEP paso
p = p + fnb (n, p) * paso
n = n + fna (n, p) * paso
NEXT ti
p0 = p
n0 = n
PRINT t, n, p
PRINT #1, t, n, p
NEXT t |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}